
Let’s go back to our copepod friend. The one who swam down to the Oxygen Minimum Zone (OMZ) to fatten up. The truth is that she only spends her days down there. In the evenings, she comes up to the surface to graze on diatoms, which are a type of algae surrounded by glass shells. Diatomaceous earth is diatom shells. With her belly full, she returns to the depths. Copepods can consume nearly their weight in diatoms in one grazing event.
Female coepopods are very anabolic. They can produce 50 or more eggs each day. If you want to be anabolic you have to close the damper. You don’t want to allow in too much oxygen or the fuel will burn away.
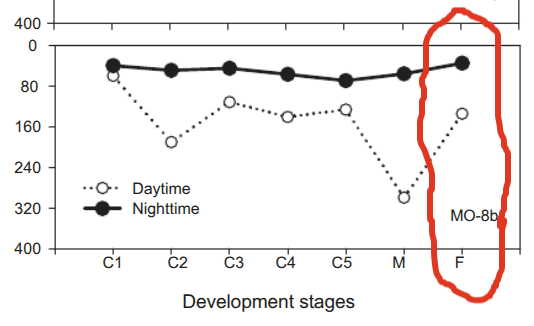
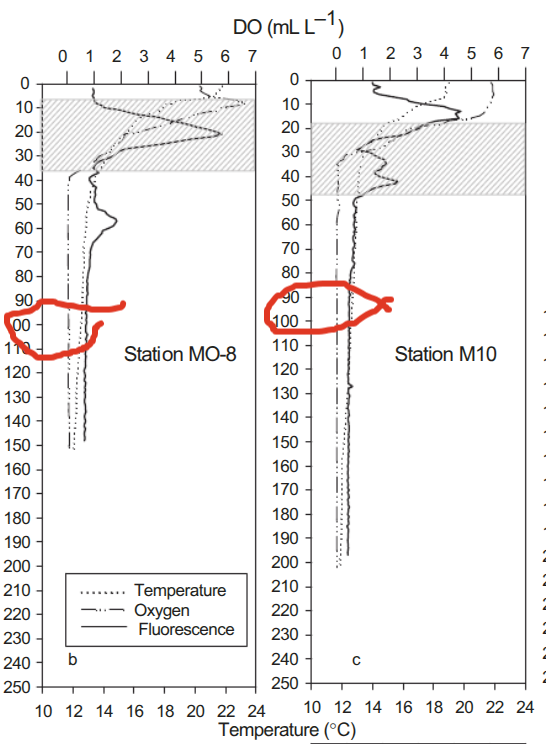
You can see that the oxygen levels are minimal while she is digesting her meal. Copepods never developed the full bag of tricks that the clever mammals (proto-mammals) did who had to live through The Great Dying on land. Then there was that asteroid that killed the dinosaurs. Ocean depths are a great buffer. The copepods just use the oxycline to get anabolic.
Copepods do another interesting thing. They convert the long chain PUFA linoleic acid and linolenic acid into very long chain (VLC) PUFA.
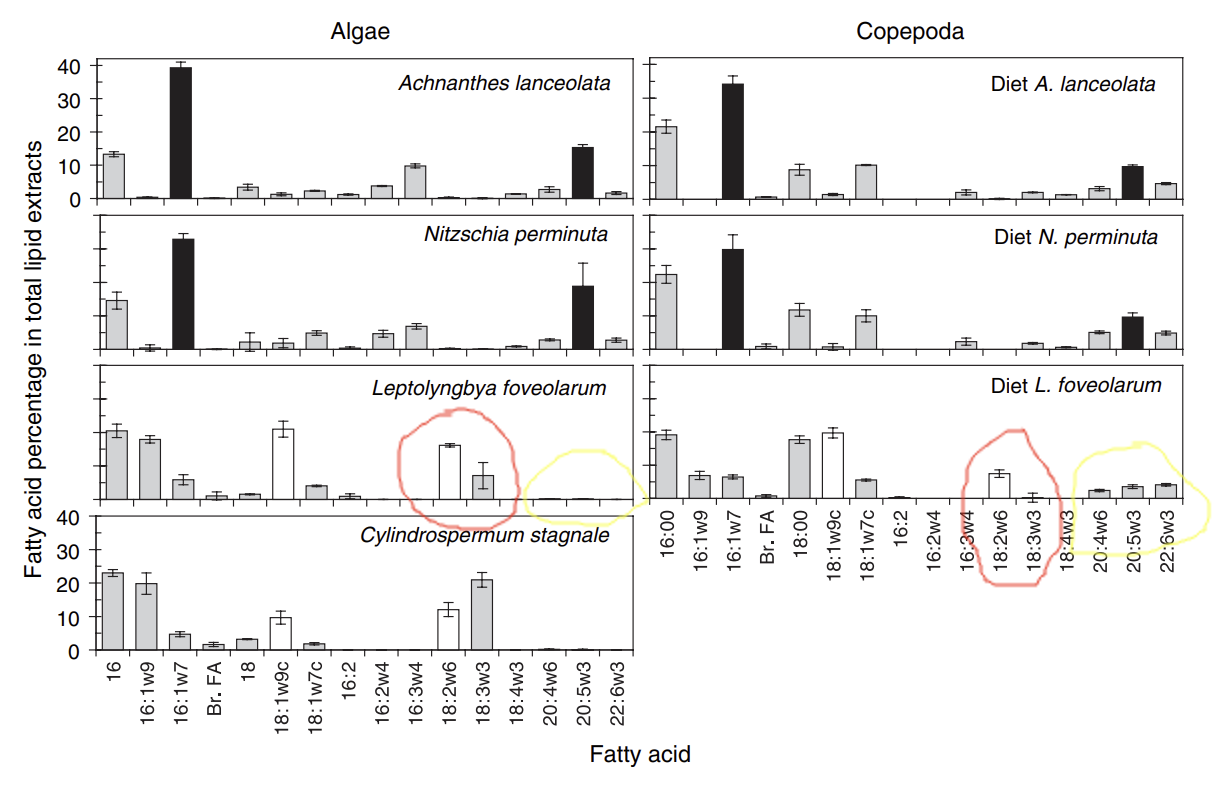
Why Copepods Desaturate and Elongate PUFA
Why do they? Let’s think about the contents of their meal. It is quite swampy, providing protein, fat and carbohydrates along with tiny shards of glass.
Protein (amino acids) is plentiful, fat is reasonably plentiful. There are carbs but they are more limited. What does the copepod need to get anabolic? She needs the building blocks for eggs: fats and amino acids. She needs NADPH with which to build. And she needs some ATP to run cellular processes. Oxygen is limiting, so we’re not expecting a ton of ATP from oxidative phosphorylation.
Here’s what she does. She runs the carbs through glycolysis to get ATP. She can get NADPH from the amino acids in an hypoxic enviroment. There is fat available. But she still has a problem.
Running glycolysis converts NAD to NADH. If the NADH builds up, glycolysis stops and she can’t get ATP. She can’t quickly regenerate NAD in her mitochondria because oxygen is limiting.
The NADH problem has afflicted life from its origins. Single celled bacteria can ferment the pyruvate produced from glycolysis to lactate and export it. This becomes more of a problem for multicelled organisms because if you dump too much lactate into the intracellular space you can die of lactic acidosis. Similar argument for fermentation to alcohol.
The animals needed a new and clever solution to the NADH problem. What else is in her belly that she can ferment to regenerate NAD. HINT: It’s not the glass shards.
D6D and D5D are enzymes whose elevation is highly correlated with metabolic syndrome in humans. Copepods have them too. Neat. They take a molecule of oxygen and a molecule of NADH. They add double bonds to PUFA. There is not a lot of oxygen in the OMZ, but there is some. The Oxygen is being redirected from the mitochondria to the oxidative fermentation of PUFA to allow ongoing glycolysis.
Fermenting the linoleic and linolenic acid gives the copepod everything it needs to continue being glycolytic and anabolic in an hypoxic world.
Did I mention that the fat cells of obese humans have very low oxygen consumption and are glycolytic? I think I did.
Caramujo, Maria‐José, et al. “Fatty Acid Profiles of Algae Mark the Development and Composition of Harpacticoid Copepods.” Freshwater Biology, vol. 53, no. 1, 2008, pp. 77–90, https://doi.org/10.1111/j.1365-2427.2007.01868.x.
Hidalgo, Pamela, et al. “Ontogenetic Vertical Distribution and Diel Migration of the Copepod Eucalanus Inermis in the Oxygen Minimum Zone off Northern Chile (20–21° S).” Journal of Plankton Research, vol. 27, no. 6, June 2005, pp. 519–29, https://doi.org/10.1093/plankt/fbi025.
Jansson, P. A., et al. “Lactate Release from the Subcutaneous Tissue in Lean and Obese Men.” Journal of Clinical Investigation, vol. 93, no. 1, Jan. 1994, pp. 240–46, https://doi.org/10.1172/JCI116951.
Kim, Wondong, et al. “Polyunsaturated Fatty Acid Desaturation Is a Mechanism for Glycolytic NAD+ Recycling.” Cell Metabolism, vol. 29, no. 4, 2019, pp. 856-870.e7, https://doi.org/10.1016/j.cmet.2018.12.023.
Peterson, William T., et al. “Diel Variations in Gut Pigment Content, Diel Vertical Migration and Estimates of Grazing Impact for Copepods in the Southern Benguela Upwelling Region in October 1987.” Journal of Plankton Research, vol. 12, no. 2, 1990, pp. 259–81, https://doi.org/10.1093/plankt/12.2.259.
Van Der Merwe, M. T., et al. “Lactate and Glycerol Release from Adipose Tissue in Lean, Obese, and Diabetic Women from South Africa1.” The Journal of Clinical Endocrinology & Metabolism, vol. 86, no. 7, 2001, pp. 3296–303, https://doi.org/10.1210/jcem.86.7.7670.
Warensjö, Eva, et al. “Associations between Estimated Fatty Acid Desaturase Activities in Serum Lipids and Adipose Tissue in Humans: Links to Obesity and Insulin Resistance.” Lipids in Health and Disease, vol. 8, no. 1, 2009, p. 37, https://doi.org/10.1186/1476-511X-8-37.